近日,我院作物病害監(jiān)測與治理團(tuán)隊胡小平課題組在Plant Biotechnology Journal期刊在線發(fā)表了題為“The wheat CRK-RLCK-MAPKs signaling module confers high-temperature all-stage resistance to stripe rust”的研究論文。該研究鑒定出一個感知條銹菌侵染與相對高溫雙重信號的關(guān)鍵分子模塊——TaCRK6-TaRLCK185-TaMAPKs,系統(tǒng)闡明了其通過調(diào)控水楊酸(SA)途徑激活小麥全生育期高溫(High-temperature all-stage, HTAS)抗條銹性的分子機(jī)制。該研究不僅為理解植物如何協(xié)同響應(yīng)生物與非生物脅迫提供了新視角,也為通過分子育種改良小麥的氣候韌性和抗病性提供了重要基因資源與理論依據(jù)。

小麥條銹病是由Puccinia striiformis Westend. f. sp.tritici Erikss.引起的典型真菌氣傳性病害,嚴(yán)重威脅小麥安全生產(chǎn)。小麥高溫全生育期抗條銹性是一種在相對較高環(huán)境溫度(18-20℃)下誘導(dǎo)產(chǎn)生的低反應(yīng)型抗性,具有非小種專化性和持久性的特點。在我國主要麥區(qū)自然條件下,該抗性通常能在條銹病病情顯著增長前被有效誘導(dǎo),是解決小麥品種抗銹性喪失問題的一條新途徑。胡小平課題組通過前期轉(zhuǎn)錄組鑒定到一個關(guān)鍵質(zhì)膜受體基因TaCRK6,進(jìn)一步解析出一條由TaCRK6-TaRLCK185-TaMAPKs構(gòu)成的線性級聯(lián)通路,通過逐級磷酸化傳遞條銹菌侵染和高溫脅迫信號,最終揭示了小偃6號高溫全生育期抗條銹性的分子機(jī)制。主要結(jié)果如下:
1.質(zhì)膜受體TaCRK6正向調(diào)控小偃6號的高溫全生育期抗條銹性過程
小偃6號在條銹菌侵染與高溫脅迫雙重作用下,TaCRK6轉(zhuǎn)錄水平顯著上調(diào)。亞細(xì)胞定位顯示,TaCRK6是一個質(zhì)膜定位的受體蛋白,推測在小偃6號感知病原體侵染與溫度脅迫的雙重信號時,質(zhì)膜定位的TaCRK6構(gòu)成識別胞外信號的“前沿哨所”。瞬時沉默TaCRK6會顯著削弱小偃6號植株的高溫抗條銹性;反之,在感病品種Fielder中過表達(dá)TaCRK6,則顯著提高植株的抗條銹性,表現(xiàn)為夏孢子堆數(shù)量減少、植物細(xì)胞壞死增加以及條銹菌生物量降低(圖1E-H)。上述試驗共同表明,TaCRK6正向調(diào)控小偃6號高溫抗條銹性過程。
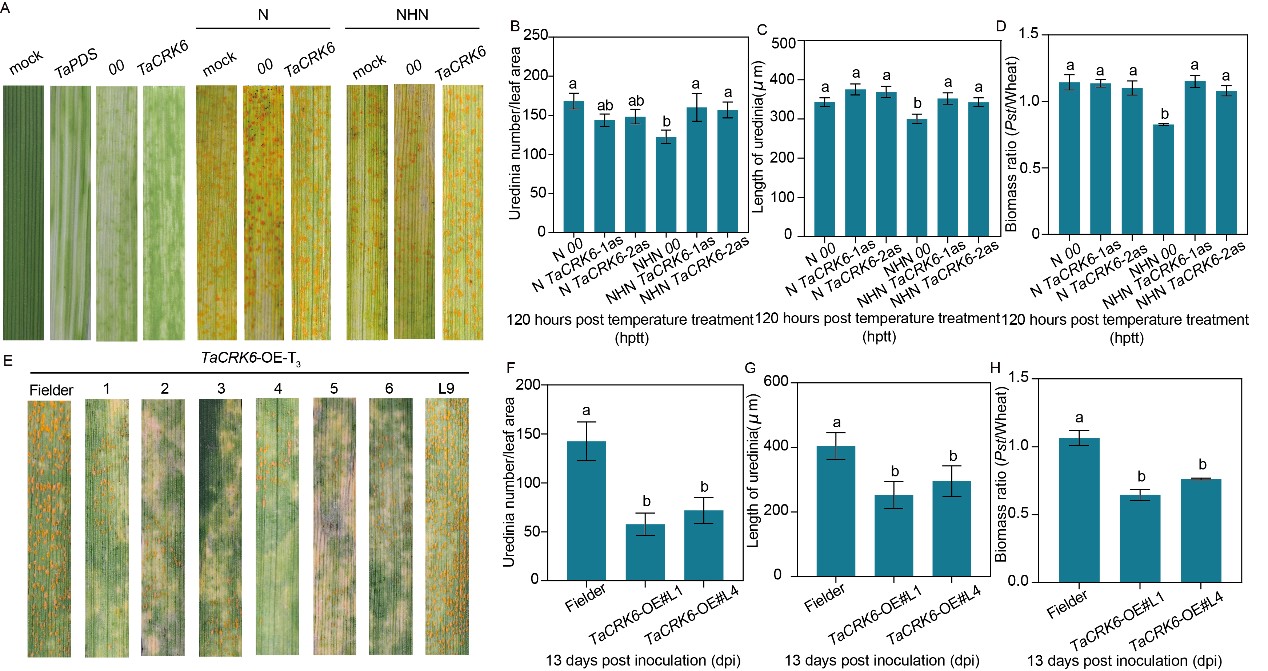
圖1 TaCRK6正向調(diào)控小偃6號小麥植株全生育期高溫抗條銹性過程
2.TaCRK6通過磷酸化TaRLCK185傳遞雙重信號,正向調(diào)控小偃6號的高溫全生育期抗條銹性過程
酵母雙雜交、雙分子熒光互補、pull-down和免疫共沉淀試驗結(jié)果均證明TaCRK6與類受體胞質(zhì)激酶TaRLCK185存在互作關(guān)系,進(jìn)一步研究發(fā)現(xiàn),TaRLCK185作為底物可被TaCRK6在體內(nèi)磷酸化。LC-MS/MS結(jié)果進(jìn)一步顯示,TaRLCK185的關(guān)鍵磷酸化位點是第248位的蘇氨酸殘基(圖2A-D)。瞬時沉默TaRLCK185顯著降低小偃6號植株的高溫抗條銹性。而在小偃6號植株中瞬時過表達(dá)TaRLCK185則顯著增強植株對條銹菌的抗性(圖2E-J)。推測TaCRK6感知雙重信號后,通過磷酸化下游TaRLCK185傳遞信號,進(jìn)而調(diào)控小偃6號全生育期高溫抗條銹性。
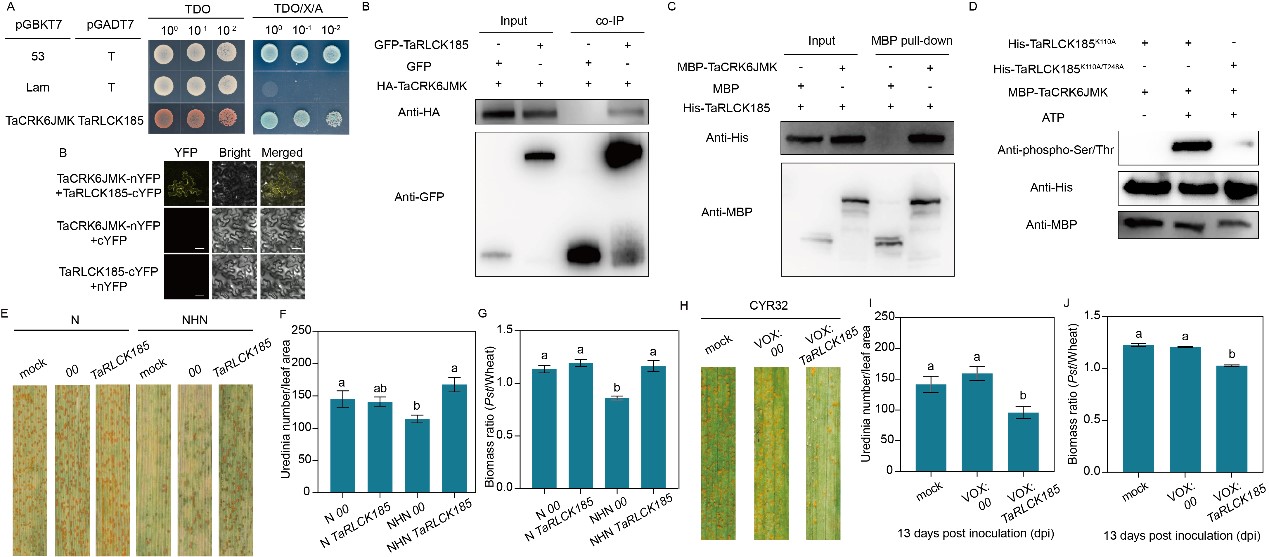
圖2 TaCRK6通過磷酸化TaRLCK185的第248位蘇氨酸殘基進(jìn)而激活小偃6號的全生育期高溫抗條銹性過程
3.TaRLCK185通過激活下游MAPK線性級聯(lián)通路正向調(diào)控小偃6號的高溫全生育期抗條銹性。
小偃6號中,接種條銹菌即可激活MAPK級聯(lián)通路的磷酸化,而條銹菌侵染和相對高溫共同處理能顯著增強植株中MAPK級聯(lián)通路的磷酸化強度(圖3A)。MBP pull-down試驗證明TaRLCK185與MAPK通路上游激酶TaMAPKKK1存在互作關(guān)系(圖3B)。同時,TaRLCK185可磷酸化TaMAPKKK1的絲氨酸和蘇氨酸殘基,該磷酸化過程依賴TaRLCK185自身激酶活性(圖3C)。此外,TaCRK6在體外可以促進(jìn)TaRLCK185與TaMAPKKK1的互作(圖3D)。在TaCRK6轉(zhuǎn)基因植株中沉默TaRLCK185部分削弱過表達(dá)植株的抗條銹性,伴隨SA通路基因TaPR1和TaPR2表達(dá)水平的抑制,夏孢子堆密度和線性長度的降低以及TaMAPKKK1轉(zhuǎn)錄水平下調(diào),說明TaCRK6-TaRLCK185-TaMAPKKK1模塊通過線性級聯(lián)調(diào)控MAPK磷酸化過程,共同介導(dǎo)小麥高溫抗條銹性過程(圖3E-J)。
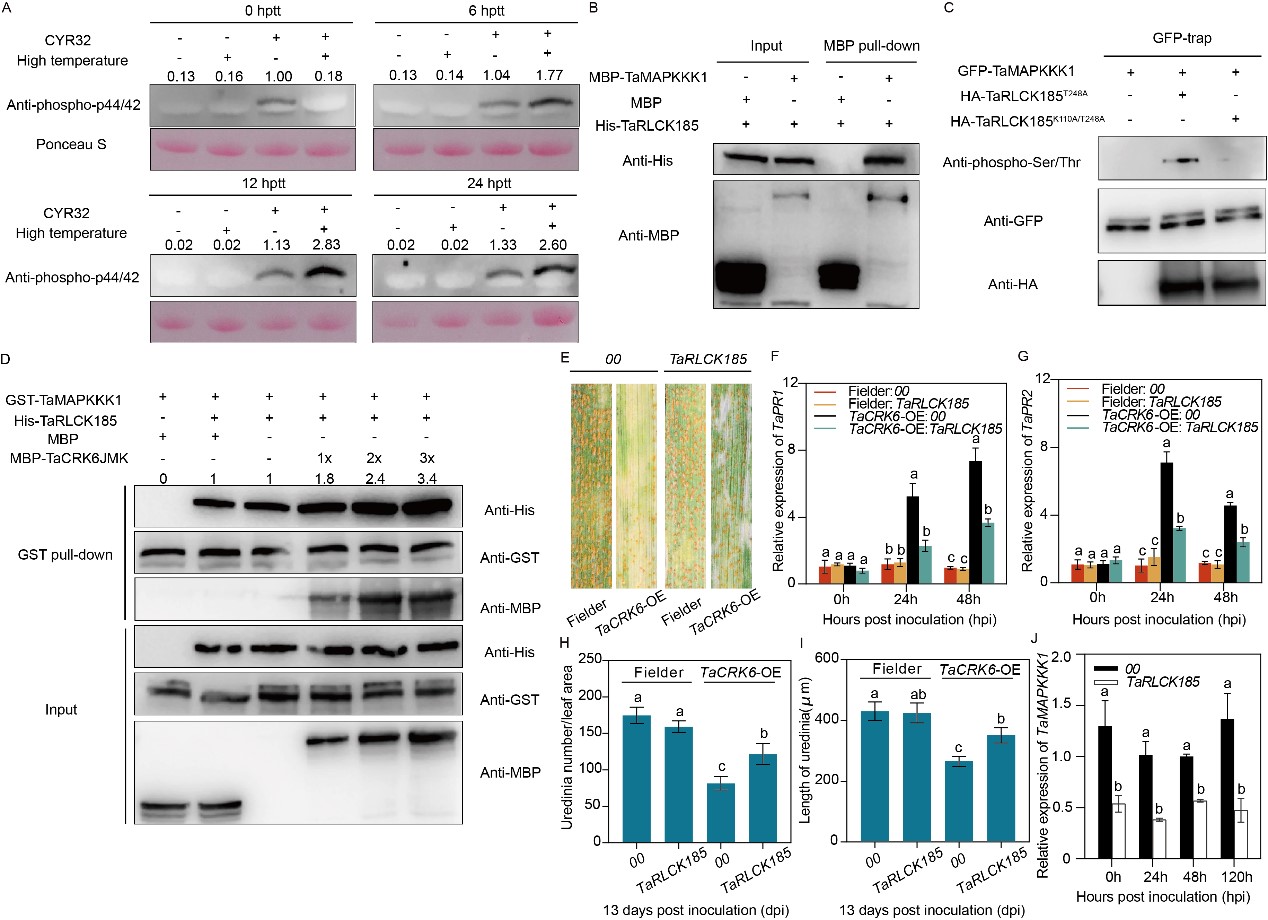
圖3 TaRLCK185通過磷酸化下游TaMAPKKK1正向調(diào)控小偃6號的高溫全生育期抗條銹性過程
4.TaCRK6-TaRLCK185-TaMAPKs模塊通過調(diào)控水楊酸信號通路介導(dǎo)小偃6號的高溫全生育期抗條銹性
研究者發(fā)現(xiàn)TaMAPKKK1與下游TaMAPKK9直接互作,而TaMAPKK9結(jié)合并激活TaMAPK6,且三者均正向調(diào)控小偃6號植株高溫全生育期抗條銹性過程(圖4A, B)。在TaCRK6沉默植株中,SA生物合成基因TaPAL的表達(dá)受到顯著抑制(圖4C)。而外源噴施SA則恢復(fù)了TaCRK6沉默植株的抗病表型(圖4D)。過表達(dá)TaCRK6則顯著提升植株內(nèi)源SA水平(圖4E)。此外,在小偃6號中瞬時沉默TaMAPK6也顯著抑制TaPAL的表達(dá)水平(圖4F)。說明該信號模塊通過調(diào)控苯丙氨酸解氨酶(PAL)途徑介導(dǎo)水楊酸生物合成過程。
該研究系統(tǒng)揭示了小麥全生育期高溫抗條銹病的信號感知與傳遞網(wǎng)絡(luò),揭示了TaCRK6-TaRLCK185-TaMAPKs核心模塊通過調(diào)控SA途徑介導(dǎo)高溫抗條銹性的新機(jī)制(圖4G)。該研究不僅為理解植物應(yīng)對多重脅迫的調(diào)控網(wǎng)絡(luò)提供了新視角,也為基于基因編輯、分子標(biāo)記輔助選擇等技術(shù)培育高產(chǎn)、抗病小麥新品種奠定了重要的理論基礎(chǔ)。
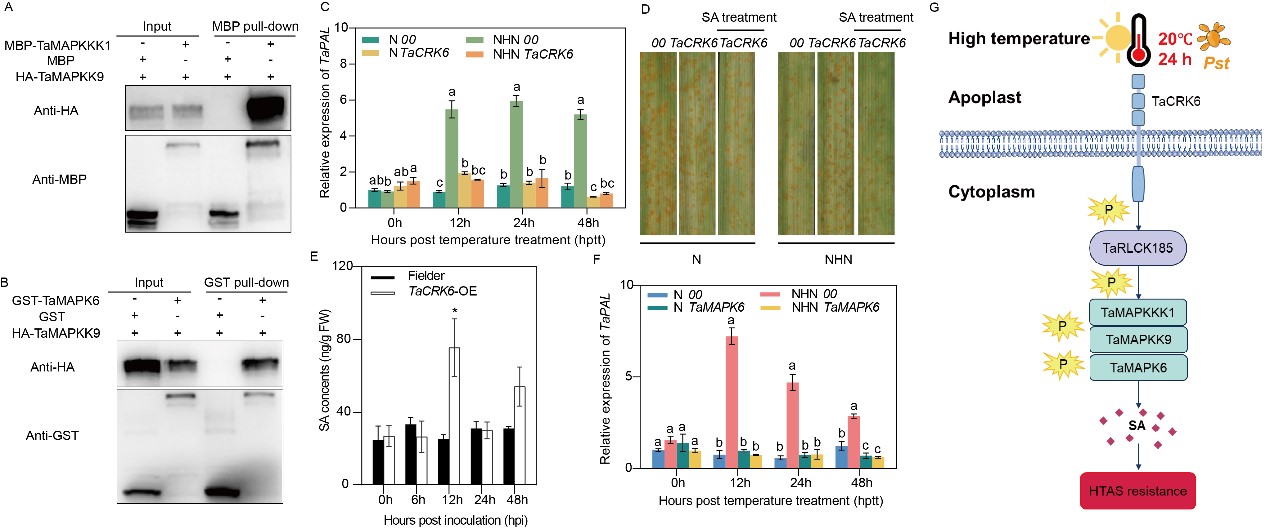
圖4 TaCRK6-TaRLCK185-TaMAPKs模塊調(diào)控小麥全生育期高溫抗條銹性分子機(jī)制
我院博士研究生師一峰為該論文第一作者,胡小平教授與李宇翔副教授為該論文共同通訊作者。該研究得到國家現(xiàn)代農(nóng)業(yè)(小麥)產(chǎn)業(yè)技術(shù)體系項目(CARS-03-37)、國家自然基金項目(13271985、31972219)、西北農(nóng)林科技大學(xué)首批學(xué)院高水平創(chuàng)新團(tuán)隊項目(XYTD2023-04)、博士后科學(xué)基金項目(2021M690130)等項目的資助。
論文鏈接:https://onlinelibrary.wiley.com/doi/10.1111/pbi.70537
編輯:劉小鳳
審核:趙 磊
